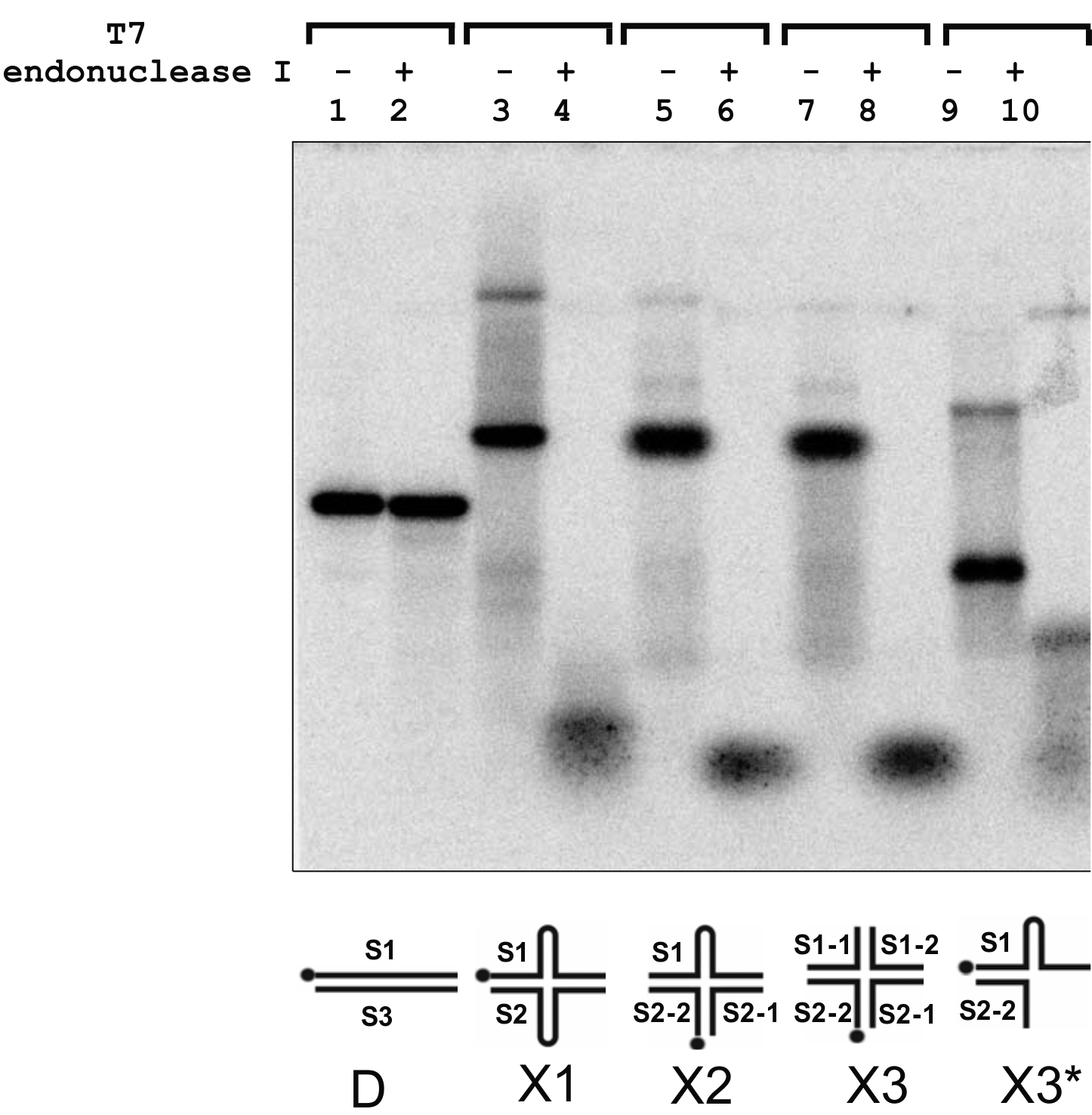
Supplementary Figure S1. Electrophoretic analysis of the cleavage products generated by T7 endonuclease I in branched DNA substrates. Digestion reactions were performed according to the manufacturer's instructions. Cleavage products were identified by electrophoresis in 7.5% polyacrylamide gel (29:1) and visualized by a phosphorimager (Perkin Elmer). Lanes 1, 3, 5, 7 and 9 - DNA substrates (25 nM) D, X1, X2, X3 and X3*, respectively, in the absence of T7 endonuclease I. Lanes 2, 4, 6, and 10 - products present in the reaction mixtures (20 μl) after incubation of a DNA substrate (25 nM) and T7 endonuclease I (0.5 unit) for 1 h at 37 °C.
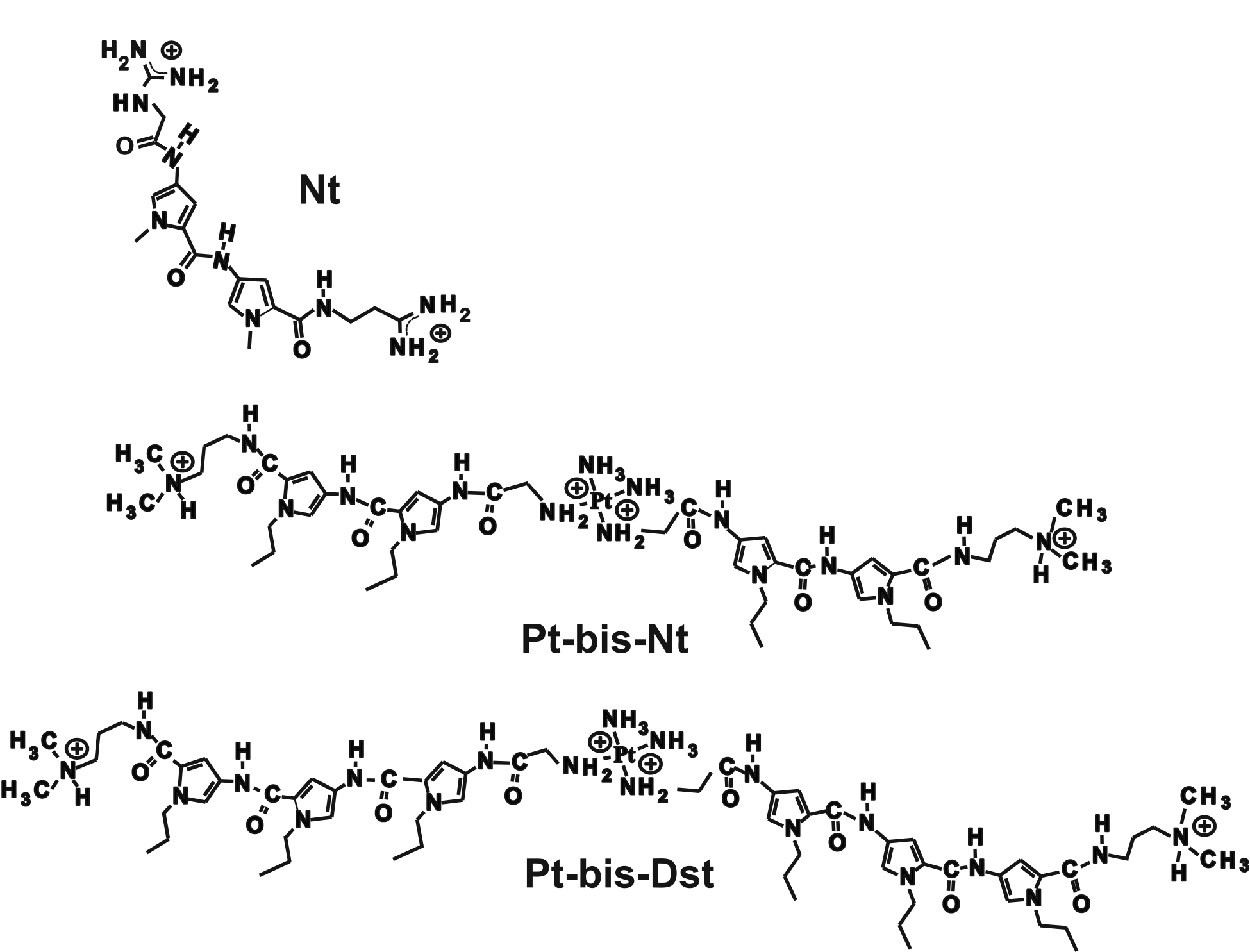
Supplementary Figure S2. Chemical structures of netropsin (Nt) and bis-linked netropsin derivatives Pt-bis-netropsin (Pt-bis-Nt) and Pt-bis-distamycin (Pt-bis-Dst).
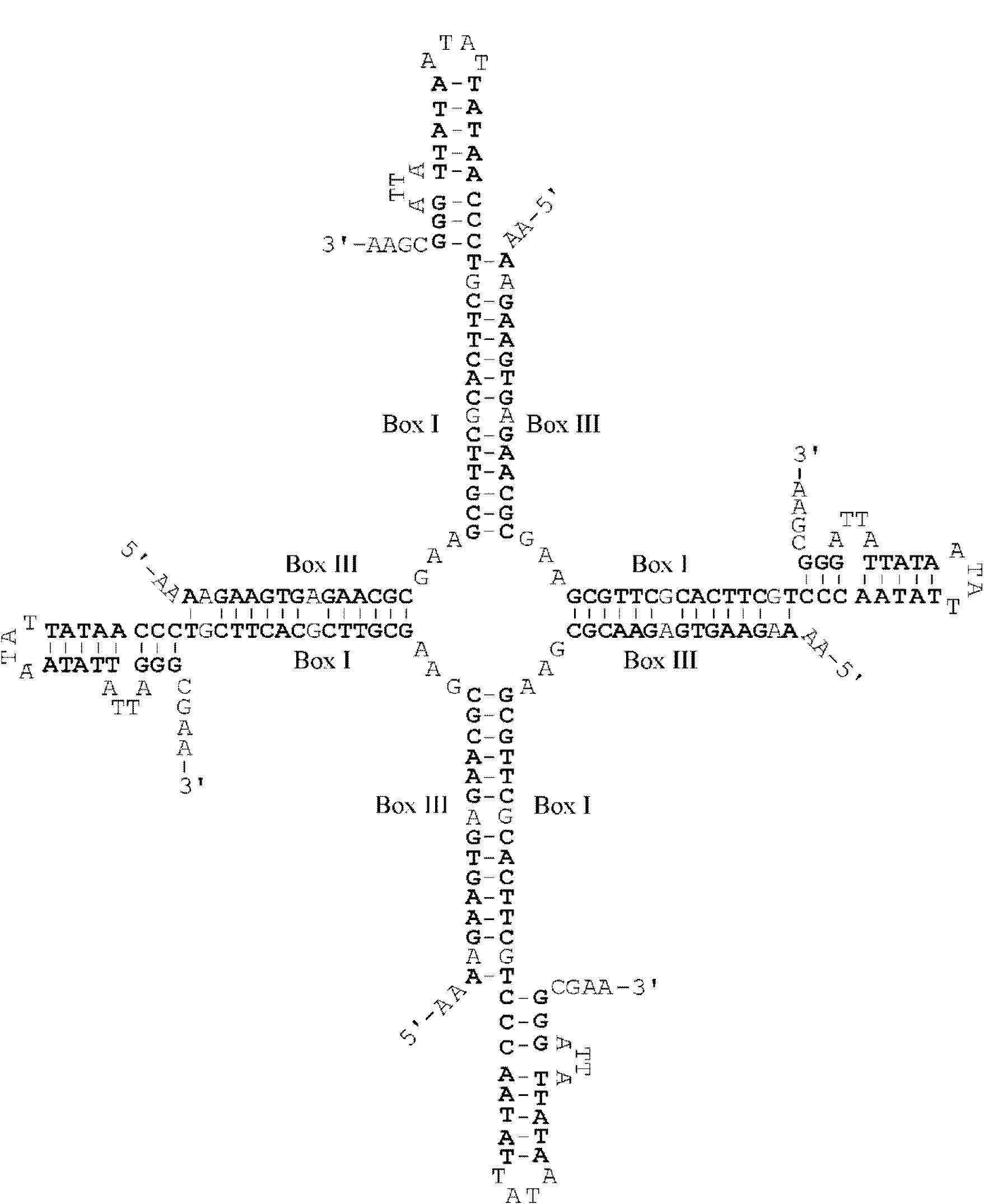
Supplementary Figure S3. The presumed model for structure of a 4-way DNA junction stabilized by complementary base pairing of the Box I and Box III inverted repeats belonging to the four V3 oligonucleotides. All the paired nucleotides are shown by bold symbols. Each arm also contains two mismatched GA pairs.
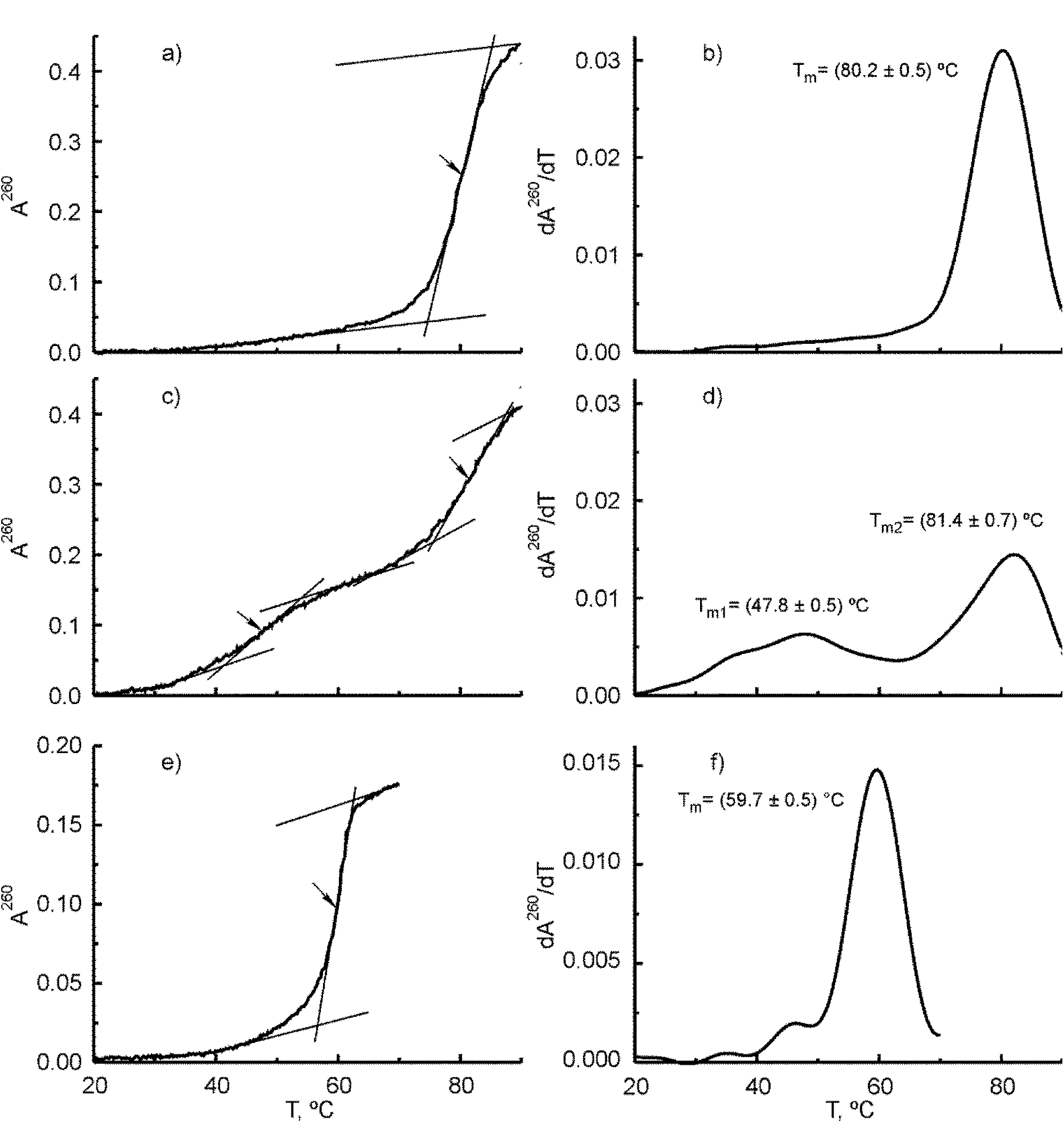
Supplementary Figure S4. Melting curves (panels a, c and e) and corresponding first derivative plots (panels b, d and f) for (V3+V4) duplex, V3 oligonucleotide and X3 junction, respectively. The melting curves were measured at 260 nm in 50 mM Tris HCl buffer (pH 8.0) containing 50 mM NaCl and 10 mM MgCl2. Concentrations of the V3 oligonucleotide, (V3+V4) duplex and X3 junction were equal to 1.76×10-6 and 8.8×10-7 and 6.1×10-7 M, respectively. A260 is the absorbance measured at 260 nm and calculated per 1 cm cell. Arrows indicate the melting temperatures for individual transitions.
In the present paper, the interactions of the origin binding protein (OBP) of herpes simplex virus type 1 (HSV1) with synthetic 4-way Holliday junctions (HJs) were studied using electrophoresis mobility shift assay and the FRET method and compared with the interactions of the protein with duplex and single-stranded DNAs. It has been found that OBP exhibits a strong preference for binding to 4-way and 3-way DNA junctions and possesses much lower affinities to duplex and single-stranded DNAs. The protein forms three types of complexes with HJs. It forms complexes I and II which are reminiscent of the tetramer and octamer complexes with 4-way junction of HJ-specific protein RuvA of Escherichia coli. The binding approaches saturation level when two OBP dimers are bound per junction. In the presence of Mg2+ ions (≥2 mM) OBP also interacts with HJ in the stacked arm form (complex III). In the presence of 5 mM ATP and 10 mM Mg2+ ions OBP catalyses processing of the HJ in which one of the annealed oligonucleotides has a 3´-terminal tail containing 20 unpaired thymine residues. The observed preference of OBP for binding to the 4-way DNA junctions provides a basis for suggestion that OBP induces large DNA structural changes upon binding to Box I and Box II sites in OriS. These changes involve the bending and partial melting of the DNA at A+T-rich spacer and also include the formation of HJ containing Box I and Box II inverted repeats and flanking DNA sequences.